


CA187 Chapt.11
Mark who is doing which.
Chapter 11: Motivational Systems of the Brain
CA188(11.1)
Hypothalamus and Eating
腦丘下部與進食
在1950年,研究者提出刺激進食由丘腦下部兩個部位來控制。下丘腦腹內側核的雙邊損傷,造成不斷地進食以及使體重增加。和其比較,下視丘外側區的雙邊損傷,造成絕食必須強迫進食才能生存下去。此項發現下丘腦腹內側核為飽足中心,抑制進食。而下視丘外側區為飢餓中心,促使進食。隨後的研究者不認同於飢餓與飽足的1950年下丘腦理論。現在認為下視丘外側區為產生一般的運動神經的損害和不靈敏的感覺中樞刺激,而不是進食明確的disruption。另外,下丘腦腹內側核為飽足中心,其有兩項重要的能力。第一為吃得過飽及肥胖為大量的下丘腦腹內側核損傷。尤其,下丘腦腹內側核損傷損害纖維物,最後終止於下丘腦的下丘腦室旁核,有些會造成纖維的通行。第二能力為吃得過飽由下丘腦腹內側核損傷產生是第二類型。下丘腦腹內側核損傷促進血液葡萄糖轉換為人體脂肪和阻止人體脂肪轉換回去葡萄糖。因此,人類下丘腦腹內側核損害吃得過飽,不是因為他們飽食中樞受到損害,而是因為當他們停止進食時,可利用的能量儲存變得危險地低。因此,下丘腦腹內側核被視為是新陳代謝中心而不是飽足中心。
相關網站:http://140.122.143.37/learning/basic/eat.htm
http://life.nthu.edu.tw/~g864264/Neuroscience/neuron/brain.htm
相關圖片:http://www.nature.com/nrd/journal/v5/n11/images/nrd2136-i1.jpg
CA190(11.2)
11.2 Subfornical Organ and Deprivation-Induced Thirst
課本內容:
水分的剝奪會以兩種不同的方式干擾身體內液體的平衡。它會產生血液容量過低,會使得血液中的鹽分濃縮,藉著細胞膜內的滲透壓把細胞內的水分驅逐出去。血液容量過低,會讓腎臟釋放腎素,這會造成血液中血管壓縮素的產生,然後這種激素作用於穹窿下器,使它產生渴覺。穹窿下器是一部分的大腦區域,位於第三腦室中間旁,就在穹窿的下面。穹窿下器對於血管壓縮素有敏感、許多的接收器,在實驗當中,偌直接把血管壓縮素注入穹窿下器,動物會感到額外的口渴。增加的滲透壓也會引起口渴,另一種滲透壓接受器位於下視丘的側視葉前區,在實驗當中,偌直接濃縮的鹽水液體注入這裡面,動物也是會感到額外的口渴。
詞彙解釋:
Subfornical organ:
位於第三腦室旁邊,穹隆的下面,對於體液的調控甚為重要,寫血管壓縮素的接收器官。
相關圖片:
http://www.endotext.org/neuroendo/neuroendo3b/figures3b/figure32.jpg
www.netterimages.com/image/5562.htm
Lateral preoptic area:
The preoptic area is a region of the hypothalamus. According to the MeSH classification, it is considered part of the anterior hypothalamus. The preoptic area is responsible for thermoregulation and receives nervous stimulation from thermoreceptors in the skin, mucous membranes and hypothalamus itself.This area propagates stimuli to either the heat-losing or heat-promoting centres of the hypothalamus.
相關圖片:
http://www.pep-web.org/document.php?id=np.001.0015.fig003.jpg
http://people.bu.edu/cshumway/images/xfvaso.JPG
CA191P11.2
CA192(11.3) 缺 劉曣晴
CA194(11.4) 缺 葉姵妏
CA196(11.5)

Pontine reticular formation是在橋腦裡的部分網狀結構,它包含膽素激性的神經核,扮演著REM睡眠的角色;也包含了正腎上腺素性和致血清素特質的神經核,扮演著SWS的角色。
Locus coeruleus是一個正腎上腺素性蛋白質的神經核,在中腦和橋腦邊緣附近,它的神經元在SWS很活躍,在REM睡眠卻很不活躍。
Raphe nuclei是延腦的縫合核,是垂直的一片致血清素特質的神經核,沿著網狀結構的中線跑,它的神經元在在SWS很活躍,在REM睡眠卻很不活躍。而人類睡眠的分期分為兩個大的時相:也就是慢波睡眠 ( Slow Wave Sleep;SWS ) 和快波睡眠(Fast Wave Sleep;FWS)。慢波睡眠又稱淺睡睡眠 ( Light Sleep )、非速眼動睡眠 ( Non-rapid-eye-movement Sleep;NREM Sleep )。
CA198(11.6)
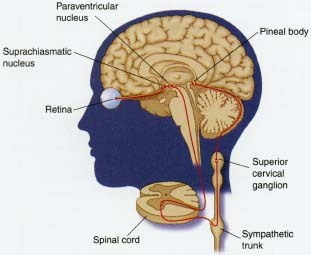
Suprachiasmatic Nucleus and Circadian Rhythms
Suprachiasmatic Nucleus
下視丘的視叉上核(suprachiasmatic nucleus)是哺乳動物的生物時鐘,控制著哺乳動物的生化、生理和行為的日週期性表現。
視叉上核有兩個,左右各一,座落在第三腦室兩旁,視神經交叉的正上方。
光照的訊息由視網膜特殊的感光細胞(節細胞)來傳遞,之後再經由視網膜下視丘徑傳到視叉上核。這條非關視覺的光訊息傳導路徑(光→節細胞(含視黑素)→下視丘的視叉上核)與視覺的光訊息傳導路徑(光→桿細胞(含視紫素)與錐細胞→節細胞→視丘→大腦 皮質)有所不同:光訊息的視覺傳導路徑讓大腦能夠分析影像,包括形狀、色彩與運動狀況;而非關視覺的光訊息傳導路徑,則是讓下視丘以及其他腦區能夠測知外界環境的光照變化,然後透過視叉上核來協調生物體內的各種活性節律。從這個角度而言,視叉上核實乃哺乳動物體內的太極,不僅獨立掌控生物體內各種陰陽變化,還能偵測天體運行(地球自轉)造成的日夜變遷,以同步協調生物體的機能運作。
視叉上核之所以這樣,是因為視叉上核神經元的細胞活性──也就是膜電特性與自發性動作電位──有著日夜不同的節律變化:白天活性高、晚上活性低。既使是在離體的狀況(單離細胞或是腦切片)也一樣。這些時鐘細胞雖然含有各種離子通道,包括鈉通道、鉀通道、鈣通道,以及其他通道,但其電活性週期變化的離子機轉卻一直是個謎。
資料來源:
http://gibms.cgu.edu.tw/gibms/modules/cjaycontent/groups/physipharm/cv/RCH.html
Circadian Rhythms
(一)生物鐘的位置:
生物體確實存在日常意義的晝夜「時鐘」,並可以告訴生物體的每日鐘點。它們的晝夜周期的誤差常常可以精確到數分鐘。研究發現具有晝夜生物鐘性質的組織按照調控機能等級可以分為:中央生物鐘(central clocks)和外周生物鐘(peripheral clocks)。從目前所知道的所有生物鐘模型研究得知,晝夜生物鐘是細胞自主的,也就是說,某些細胞就具有生物鐘的特性。雖然晝夜生物鐘受外界環境(比如光照溫度等條件)的調控,複雜生物的中央晝夜生物鐘往往並不存在於光感受器上。比如,哺乳動物的中央晝夜生物鐘存在於下丘腦的視交叉上核(Suprachiasmatic nucleus, SCN)。
(二)生物節律的種類:
1.超晝夜的(亞日的)節律(Infradian Rhythmus)
該詞源於拉丁語:「infra」為「底下」,「dies」為「日」,亦即周期比一天長的節律。 例如鳥類的遷徙;季節性的(大概 365.25天長)冬眠;還有與退漲潮相關的半月周期,如在滿月、新月出現大潮,而半月時出現小潮(大概 14.25 天),銀漢魚只在漲潮時在岸上產卵;或者太陰日節律的,以28.5為周期(磯沙蠶屬)。
2.近潮汐節律(Circatidal Rhythmus)
跟隨12.5小時的潮汐節律。一些海岸線的動物有這種節律,例如水生的蟹類動物漲潮時才會活動,而生長在岸上的蟹則會在退潮時覓食。
3.次晝夜(超日)的節律(Ultradian Rhythmus)
源於拉丁語的「ultra」(超)和「dies」(天、日),其頻率超過日頻率,就是說一天出現兩次以上(嚴格來說是整數次,這是與近潮汐節律的區別)。這些短於24小時的節律的例子有蝙蝠的捕食周期、成人90分鐘睡眠循環、垂體的間歇性荷爾蒙分泌等。
4.近晝夜節律(Circadiane Rhythmus)
來自拉丁語「circa」(大約)和「dies」,為接近24小時長的節律,如人類睡眠和蘇醒、植物的葉運動等。
(三)單細胞生物:
生物鐘的運行並不一定需要一個網路作為硬體。
藻類如眼蟲屬或衣滴蟲有趨光性晝夜節律。
草履蟲有晝夜生理過程。
海生的腰鞭毛蟲, 如多邊膝溝藻,也有自己的晝夜節律。它在日出前一個小時就會浮到水面,形成厚厚的一片,進行光合作用。在有利條件下它們會形成紅潮。在日落之前它們則會重新潛到海中。晚間它們借助熒光素酶發出生物光,人們推測這是可以驅趕天敵撓足亞綱的。 這些節律也可以在實驗室里通過施加持續的影響而發生。
同時原核生物 (細菌,和藍藻)也有晝夜節律。
(四)植物;
在植物中仍沒找到生物鐘的中央控制部分或是起搏點。『現在只能推測,光合作用以及與之聯繫的運動時由遍佈植物體的多個時鐘共同控制的』。
例如光合作用器官的新陳代謝,在實驗中可以觀察到是由於光照對基因表達產生影響引起的。 每天在葉綠體的類囊體膜上的光收集器(Lhc)都會進行光合作用。光會影響細胞核基因的轉錄和翻譯。西紅柿到目前為止已發現19個Lhc-基因。
目前在Lhc-基因的運作機制和其啟動子方面進行著很多的研究。
(五)動物:
在動物中起搏點位於中樞神經系統。對於昆蟲如果蠅存在腦部的腹側的側邊小神經元(Small Ventral Lateral Neurons, sLNv)中,這些神經元表達色素擴散因子(Pigment Dispersing Factor, PDF)。不在光葉中。對於軟體動物在視網膜的基底部對於脊椎動物在視交叉上核和松果體(Pineal organ,Epiphyse)中。松果體分泌褪黑激素(N-乙醘-5-甲氧基色胺)。魚,兩棲類動物,爬行類動物和很多鳥類動物中松果體是對光敏感的,除此之外它還控制了除褪黑激素晝夜產生節律外的其他節律,如體溫和進食。從中可得知,松果體比視交叉上核更早掌管著生物節律。
(六)哺乳類動物:
哺乳類動物中松果體和視交叉上核共同控制了節律,但還有很多其他證據表明,還有其他起搏點的存在,如視網膜
(七)人類:
生物鐘對我們的重要性
第一:
我們的生活模式越來越偏離生物鐘。輪班制越來越多。
第二:
我們越來越少去曬太陽。特別在冬天,我們在室內過上大部分的時間,光強度鮮有高於500流明。在戶外即使是陰天最少有8000流明,而太陽光則有100000流明。因此就生物鐘系統來說我們大多生活在黑暗中。我們的晝夜節律其實每天都需要一次新的「校正」,但現在卻遇上了很大的困難。後果可能是失眠和飲食失調,精力不足直到深度抑鬱症。在北歐(如挪威), 在冬天光效率甚至直逼0。在當地,為治療冬天抑鬱症人們採取了光療法。
第三:
我們越來越頻繁的跨時區旅遊(即從東向西,或從西向東),這是對我們晝夜節律一個重大挑戰。
時間利用的習慣分成兩類。
一類晚睡晚起,睡眠時間長——「貓頭鷹型」,而「雲雀型」則是早睡早起。這個差別是基因素因引起的,所以要改過來是很難的。這也意味著,我們大部分人是逆節律生活的。 青春期年輕人幾乎全是貓頭鷹型,因此推遲上課時間一個小時,特別是在冬天,無論對授課效果還是健康都是大有好處的。 除了這兩種類型外,還有睡眠時間長短之分。這些類型可以相互組合。還有一種類型的人,他們對睡眠和日光同步束手無策。
生物鐘學與我們的年齡有關。
嬰兒時期次晝夜系統(短的活動時間)和長的睡眠交替,直到晝夜系統發展到能夠掌管生物鐘為止。但隨著年齡的增長它也會漸漸失效。這也是老年人睡眠和活動障礙的原因。
資料來源:
http://zh.wikipedia.org/w/index.php?title=%E6%99%82%E9%96%93%E7%94%9F%E7%89%A9%E5%AD%B8&variant=zh-tw


CA200(11.7)

終腦:前腦兩部分的其中之ㄧ,另一部分為間腦;間腦:腦幹的最前區,在終腦和中腦之間;中腦:在後腦和間腦之間,為腦幹的一部分;後腦:終腦和隨腦之間,屬後腦(hindbrain)的一部分(另一部分為隨腦);隨腦:大腦最後部份,在腦幹中後腦和脊椎之間
CA201C 11.7

{kind=link}
{kind=link}
{kind=link}
{kind=link}
ventromedial nucleus:控制女性性行為的主要細胞核;periaqueductal gray:中腦中的周圍大腦導管部位;medial preoptic area:控制男性性行為主要的細胞核;lateral tegmental:控制男性性行為的經由腹部中央核傳遞的路徑;sexually dimorphic nucleus:medial preoptic area的細胞核,在公鼠腦中的比在母鼠腦中的大
CA202(11.8)

suprachiasmatic nucleus
下視丘的視叉上核是哺乳動物的生物時鐘,控制著哺乳動物的生化、生理和行為的日週期性表現。視叉上核有兩個,左右各一,座落在第三腦室兩旁,視神經交叉的正上方。光照的訊息由視網膜特殊的感光細胞(節細胞)來傳遞,之後再經由視網膜下視丘徑傳到視叉上核。這條非關視覺的光訊息傳導路徑(光→節細胞(含視黑素)→下視丘的視叉上核)與視覺的光訊息傳導路徑(光→桿細胞(含視紫素)與錐細胞→節細胞→視丘→大腦 皮質)有所不同:光訊息的視覺傳導路徑讓大腦能夠分析影像,包括形狀、色彩與運動狀況;而非關視覺的光訊息傳導路徑,則是讓下視丘以及其他腦區能夠測知外界環境的光照變化,然後透過視叉上核來協調生物體內的各種活性節律。從這個角度而言,視叉上核實乃哺乳動物體內的太極,不僅獨立掌控生物體內各種陰陽變化,還能偵測天體運行(地球自轉)造成的日夜變遷,以同步協調生物體的機能運作。
視叉上核之所以有這種能耐,是因為視叉上核神經元的細胞活性──也就是膜電特性與自發性動作電位──有著日夜不同的節律變化:白天活性高、晚上活性低。既使是在離體的狀況(單離細胞或是腦切片)也一樣。這些時鐘細胞雖然含有各種離子通道,包括鈉通道、鉀通道、鈣通道,以及其他通道,但其電活性週期變化的離子機轉卻一直是個謎。一直要到最近,才有人發現到鈣通道與細胞內鈣離子釋放的節律變化或許與這些細胞的活性變化有關。
medial preoptic area視前內側區,接受NE系統影響並參與冬眠調製的主要部位
paraventricular nucleus下丘腦室旁核分泌催產素
lateral hypothalamus 外側下丘腦
ventromedial nucleus下丘腦腹內側核
下丘腦是中樞神經系統中調節攝食和能量代謝的主要部位。下丘腦外側部可能存在“攝食中樞”,電刺激此區引起攝食增加,損毀此區導致動物攝食減少或拒食。下丘腦腹內側核可能存在“飽中樞”,電刺激此區引起動物攝食減少,損毀此區導致動物攝食增加而發胖。動物在飢餓或血糖水平降低時,下丘腦外側區神經元活動增加,腹內側核神經元活動減弱;靜脈注入葡萄糖後,下丘腦腹內側核神經元活動增加,下丘腦外側區神經元活動減弱。可見兩中樞神經元活動有交互抑制現象。
No comments:
Post a Comment